Our Programs
Science
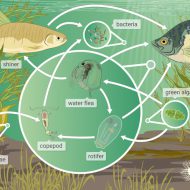
The LWRC’s Science Program facilitates whole ecosystem research and monitoring on Lake Winnipeg through its on-lake infrastructure and annual meetings.
Learn More
Education
Providing unique experiential learning opportunities for schools over a wide range of grades
Learn More
Scholarship
Honours and Graduate Student Scholarship offered to encourage and promote research initiatives by young scientists on Lake Winnipeg.
Learn More